
GOMEL STATE MEDICAL UNIVERSITY
Normal and Pathological Physiology Department
PHYSIOLOGY OF EXCITABLE TISSUES
Physiology of nerve tissue
Lecturer:
Victor Melnik
Professor,
Doctor of Biological Sciences

Lecture plan:
1. Membrane-ionic theory of the origin of the resting
membrane potential.
2. Membrane action potential (AP). Changes of
excitability during excitation. Laws of stimulation and
assessment of excitability. Lability.
3. Physiology of nerve fiber. Laws of excitement
conduction. Mechanisms of signal formation and
conduction in myelinated and unmyelinated fibers.
4. Parabiosis.
5. Physiology of synapses. Mechanisms of signal
transmission in chemical synapses. Principles and
features of excitation transmission in interneuronic
synapses.
6. Perception of external stimuli (reception).
Transformation of stimulus energy.

1. Membrane-ionic theory of the origin of the resting
membrane potential
All tissues are excitable, but conventionally they are divided
into excitable and non-excitable. Nervous, muscular, and
glandular tissues are excitable, as impulses which appear in
them go along the membrane. These impulses have an
important diagnostic value (for example, in
electrocardiography, electroencephalography,
electromyography, etc.).
The cell membrane is known to have an electric charge. Its
external surface is charged positively ―+‖ and the internal
one — negatively ―–―.
The difference between the charges of the external and
internal membrane sides is called the resting membrane
potential.

FIGURE — CHARGES DISTRIBUTION
BETWEEN INSIDE AND OUTSIDE OF THE CELL
IN ITS RESTING STATE

FIGURE — MEASUREMENT OF THE MEMBRANE POTENTIAL
OF THE NERVE FIBER USING A MICROELECTRODE

The formation of the resting membrane potential
(RMP) depends on the concentrations of К
+
, Nа
+
,
Са
2+
, Сl
-
, as well as on the features of the cell
membrane. The cell membrane has 3 layers (Figure):
External layer — mucopolysaccharides.
Bimolecular lipid layer.
Internal (protein) layer.
The membrane has channels which have the
properties of:
Selectivity — the channels are divided into 4
groups: sodium, potassium, calcium, chloric.
Selectivity is not obligatory yet preferable.
Electroexcitability.

FIGURE — STRUCTURE OF THE PLASMA
MEMBRANE

FIGURE — SODIUM AND POTASSIUM
CHANNELS
Many
channels can
be opened or
closed by gates
that are
regulated by
electrical
signals or
chemicals that
bind to the
channels. The
gating of protein
channels
provides a
means to
control their ion
permeability
(Figure).

Classification of ion channels:
By the amount of ions to which the channel
is permeable:
— Selective ion channels (permeable to one
type of ions).
— Non-selective ion channels (permeable to
several types of ions).
By the type of ions the selective channels are
divided into K
+
, Nа
+
, Са
2+
, Сl
-
channels.

By the type of regulation (gating):
— Voltage-gated channels. They react to the
changes of the membrane potential. When the potential
reaches a certain value, the channel becomes activated
and ions pass through it down the concentration
gradient.
— Chemically-gated channels (ligand-gated
channels). In these channels the gates are opened by
the binding of a chemical substance (a ligand) with
receptors.
— Mechanically-gated channels. In these channels
the permeability is changed if there are some
mechanical actions on the membrane (these channels
are present in the membrane of the mechanoreceptors
of the blood vessels, skin, etc).

In cells at rest all sodium channels are closed.
There are leakage channels (non-specific), which are
permeable to all elements but are most permeable to
potassium. They are always open, and potassium ions
move through these channels down the concentration
and electrochemical gradients. According to the
membrane-ionic theory, the presence of the RMP is
caused by:
Unequal ion concentration inside and outside the
cell.
Different permeability of the channels to these ions.
There are many K
+
ions inside cells and few outside
them, opposite to Nа
+
. There are slightly more Сl
-
ions
outside cells than inside them. There are a great
number of organic anions inside cells.
The membrane of cells at rest is only
permeable to K
+
ions. At rest, potassium
ions constantly move outside cells, where
there is a high Nа
+
concentration.
Therefore, in cells at rest, the external
surface of the membrane is positively
charged. High-molecular organic
anions (proteins) are concentrated on
the internal surface of the membrane and
determine its negative charge. Due to
electrostatics they keep K
+
ions on the
other side of the membrane. The basic
role in the formation of the RMP
belongs to K
+
ions (Figure).

FIGURE — IONIC MECHANISM OF THE FORMATION
OF THE RESTING MEMBRANE POTENTIAL

Despite the streams of ions coming through the
leakage channels, the ion concentrations are not
equivalent, i. e. they are always constant. This
does not happen because of the existence of Nа
+
-
K
+
-pumps in the membranes (Figure).
They continuously pump Nа
+
out of cells and
pump K
+
against the concentration gradient into
the cytoplasm. For 3 Nа
+
ions removed from a cell,
2 K
+
ions are introduced into it. The transmission of
ions against the concentration gradient is carried
out by active transport (with energy input).

FIGURE — STRUCTURE OF THE SODIUM-
POTASSIUM PUMP

Membrane potentials in different tissues
are characterized by different values: the
highest one is in muscular tissue —
80–90 mV, in nervous — 70 mV, in
connective — 35–40 mV, in epithelial
— 20 mV.
When the internal charge of the
membrane becomes less negative, it is
known as membrane depolarization. If
the internal charge of the membrane
becomes more negative, it is called
hyperpolarization.

2. Membrane action potential (AP)
Being imposed by a threshold stimulus, the
permeability of the membrane changes, and an
action potential (AP) or excitation occurs (Figure).
AP is rapid fluctuations of the membrane
potential during excitation.
The threshold stimulus is the minimal strength
which leads to the minimal response. To characterize
the threshold stimulus, the concept of rheobase (in
Greek, the root rhe translates to "current or flow", and
basi means "bottom or foundation") is used.
Apart from the threshold stimulus, there are
subthreshold stimuli which cannot generate
responses but induce a shift in cell metabolism.
Besides, there are superthreshold stimuli.

Having arisen, AP goes along the membrane
without changing its amplitude. It has the
following phases:
1. Slow depolarization (See figure «2»);
2. Fast depolarization (See figure «3»).
3. Fast repolarization (See figure «4»);
4. Slow repolarization or negative
afterpotential (See figure «5»).
5. Hyperpolarization or positive
afterpotential (See figure «6»).
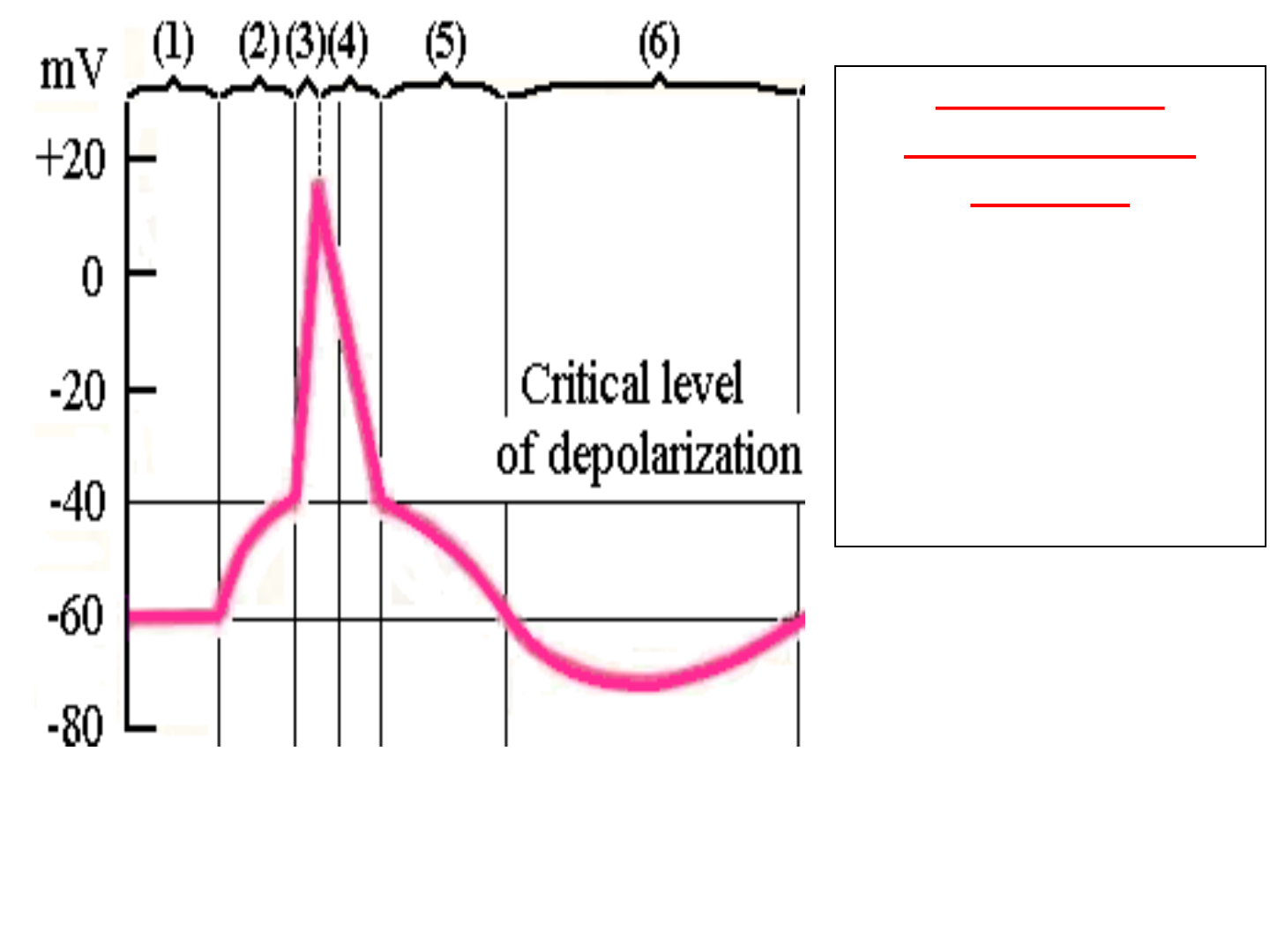
FIGURE — MEMBRANE ACTION POTENTIAL
Phases of the
membrane action
potential:
(
2) Slow depolarization.
(
3) Fast depolarization.
(
4) Fast repolarization.
(
5) Slow repolarization.
(
6) Hyperpolarization.

Mechanism of the AP origin (Figure).
Under the effect of the threshold stimulus,
the cell membrane becomes permeable to
Na
+
ions, which stream inside the cell at a
high speed (the flow of Na
+
ions into cells is
higher than the flow of K
+
ions outside cells).
The internal side of the membrane
becomes positive, and on its surface a
negative charge is formed. The changes of
the charges on the internal and external
surfaces of the membrane correspond to the
depolarization phase (See figure «1»).

Afterwards the sodium channels
close, and the potassium channels
which have been partially closed open.
K
+
ions go out of the cell. This AP
phase is called repolarization (See
figure «2»)
The action of the Nа
+
-K
+
-pump and
the RMP are restored (See figure «3»).
The basic role in the formation of
AP belongs to Na
+
ions.

FIGURE — SUCCESSIVE
OPENING AND CLOSURE OF CHANNELS
I. Sodium
channel
1. Opening
2. Closure
3. Opening
II. Potassium
channel

Changes of excitability during excitation
During the development of AP (excitation), the
excitability of cells changes (Figure).
The development of the slow depolarization phase
raises the excitability (hyperexcitability) creating
conditions for a response. Further, when the slow
depolarization phase is replaced by the fast one, the
excitability rapidly reduces and, when the
repolarization phase occurs, it starts to recover
again.
There are several periods of excitability:
1. Refractory period:
а) absolute;
b) relative.
2. Supernormal or exaltation period.

The refractory period is an interval of time during
which a cell cannot respond to the action of a stimulus.
The sodium channels are inactivated. During the
absolute refractory period, the cell does not respond to
the action of threshold or superthreshold stimuli.
The membrane repolarization leads to the reactivation
of the sodium channels. This is the relative refractory
period. During this period, a response may appear
under the action of the superthreshold stimulus.
During the supernormal period, excitability exceeds
the initial level. At this state the cell can respond to a
stimulus the strength of which is a bit lower than the
threshold one. The threshold of excitation is decreased
because the values of the membrane potential are close
to the critical level.

FIGURE — CHANGES OF THE MEMBRANE POTENTIAL
AND EXCITABILITY DURING EXCITATION
I — Changes of
the membrane
potential:
(1) Membrane
resting potential
Phases of
membrane action
potential:
(2) Slow
depolarization;
(3) Fast
depolarization.
(4) Fast
repolarization;
(5) Slow
repolarization
(6) Hyperpolar-
tion
II — Changes
of excitability:
(a) Normal
excitability
(b) Absolute
refractory period
(c) Relative
refractory period
(d) Supernormal
or exaltation
period

Laws of stimulation and assessment of excitability.
Lability.
The excitability of tissue depends on the threshold of its
irritability (rheobase). Rheobase is the minimal strength of a
stimulus that is able to cause excitation of tissue and induce
the minimal response. The lower the strength of the threshold
stimulus is, the higher the excitability of tissue is. However,
the response of tissue depends on the strength of the stimulus
to a certain extent.
The response of the cell also depends on the duration for
which the stimulus is applied. The threshold strength of the
stimulus is in the inverse relation with its duration.
The interrelations between the strength and duration of the
stimulus are demonstrated by the strength-duration curve. If
the strength of the current is «1», to induce a response from
tissue, the duration of the stimulus must be «а» (Figure).

Figure — The strength-duration
curve
Notes: 1 — rheobase; 2 — double rheobase;
a — useful time; b — chronaxie

The shortest duration for which a stimulus equal
to rheobase should react to induce a response is
called the useful time (Figure). If to double the
strength of the stimulus («2» — two rheobases), the
duration of the stimulus necessary to induce the
response decreases («b») (Figure).
The shortest duration for which a stimulus equal
to double rheobase should be applied on tissue to
cause a response is called chronaxie.
If the strength of a stimulus is equal to half of
rheobase (half of «1»), no response will arise
regardless of the duration of the stimulus. For
example, the reflex of withdrawing hands away from
a cold iron will not occur.

If tissue is exposed to a stimulus whose
strength is equal to triple rheobase, but whose
duration is too short (half of «b»), no response
will arise either. For example, if to touch a hot
iron very quickly, it is impossible to feel its
temperature (Figure).
Chronaxie characterizes the rate of excitation
generation. In different tissues it varies, which is
used for medical purposes, e.g. to determine the
damage of motor nerves.
.
Lability
To characterize the development of separate
APs, the concept of lability is used. Lability is the
rate of the development of the response to a
stimulus (separate APs). The higher lability is,
the more APs tissue can make per unit of time.
The measure of lability is the highest number of
stimuli to which tissue can respond by generating
APs per unit of time. The maximal rhythm of
excitation is limited by the duration of the absolute
refractory period. If the refractory period lasts for
0.5 msec, the maximal rhythm is 1,000 impulses
per second and more.

Nervous tissue possesses the highest
lability. It can generate up to 1,000
impulses per second. Muscular tissue can
conduct up to 500 impulses per second.
Synapses are least labile. However,
tissues cannot function at the maximum
rhythm for a long time. In natural
conditions tissue reacts to the excitation of
a lower rhythm which can be kept for a
long time. This rhythm is produced during
the supernormal period and is therefore
called optimal. In nerve fiber it is 500
impulses per second, in muscle fiber —
200 impulses per second.

During rhythmic excitation, lability can
increase or decrease. Decreased lability leads to
the development of the processes of inhibition
and its increase determines the properties of
tissues to adjust to a new higher rhythm of
impulses. The adjustment to the higher rhythm
is connected with the pumping of Nа
+
ions out
from the cytoplasm during excitation. Thus,
muscles are capable to adjust to a more frequent
rhythm of impulses coming to them from nerve
fibers. For example, if after a long flight you see
your parents at the airport, your tiredness
disappears for a while. This is connected with
the adjustment of your muscles to a higher
rhythm coming from the nerve centers.

FIGURE — LABILITY OF VARIOUS TISSUES
Synapses Muscles Nervous tissue

3. Physiology of nerve fiber
Nerves specialize on the conduction of
stimuli and connect the nerve centers
with executing organs. Nerves consist of
myelinated and unmyelinated fibers,
which are coated with the connective
tissue membrane (Tablе).
The surface of the axial cylinder of
nerve fiber is coated with the plasma
membrane, which performs the main
role in the generation and conduction of
excitement.

Table — Properties of different nerve fibers
Type
of fibers
Diameter,
mcm
Speed of
conduction,
m/sec
Functions
Aα
(myelinated)
13–22 70–120
Efferent
fibers conduct
excitation
to
skeletal muscles, afferent
fibers
conduct
excitation from
muscle
receptors
Aβ
(myelinated)
8–13 40–70
Afferent
fibers conduct
excitation
from
touch and
tendinous
receptors
Aγ
(myelinated)
4–8 15–40
Afferent
fibers conduct
excitation
from
touch and pressure
receptors,
efferent
fibers conduct excitation
to
skeletal
spindles
B
(myelinated)
1–3 3–14
Preganglionic
fibers of
the
vegetative
nervous system
C
(unmyelinate
d)
0.5–1.0 0.5–1.0
Postganglionic
fibers of
the
vegetative
nervous
system,
afferent
fibers conduct
excitation
from
pain, temperature,
and
pressure
receptors

Myelinated fibers have an intercept sheath,
which is formed by myelin segments 1–2 mm long
(the myelin sheath). The gap between the two
segments is called the node of Ranvier (Figure).
Figure — Structure of myelinated fiber

The myelin sheath is deposited around the
axon by Schwann cells. The membrane of
Schwann cells first envelops the axon. Then
Schwann cells rotate around the axon many
times, laying down the multiple layers of the
Schwann cell membrane. Myelin is highly
resistant and besides it performs the isolating
function and takes part in the metabolism of
nerve fibers. A signal along myelinated fiber
goes only through the nodes of Ranvier, as
they have many sodium channels.

Unmyelinated fibers are of the similar structure
but have no myelin. Their surface is coated with
Schwann cells.
If to dissect nerve fiber, its peripheral end after a
while loses the ability to conduct signals and
degenerates. Myelin undergoes fatty degeneration
and transforms into fatty drops. The central end of
nerve fiber is able to regenerate. A growth bulb is
formed on it and grows towards the periphery (from
0.4 to 4.5 mm a day) and reaches the
corresponding organ or tissue. Therefore, their
innervations are recovered. Thus, the first signs of
the regeneration of muscle innervations can appear
after 5–6 weeks.

Laws of excitement conduction
• The anatomical and physiological integrity of
fibers is essential. Dissection or compression
affects the conductivity of nerves. If to cut a nerve
and separate both the ends of the cut at a distance
of 1 mm, excitation can skip from one end to the
other only through myelinated fibers.
• Signals may propagate along nerves in both
the directions. This law is typical only for fibers
isolated from the body, as inside the body signals
are transmitted through synapses which conduct
APs only in one direction.
• Isolated signal conduction, i.e. a signal from one
nerve fiber cannot skip to another one located in
parallel.

Mechanisms of signal formation and
conduction in myelinated and
unmyelinated fibers
The mechanism of signal conduction
in unmyelinated fibers. The action of the
threshold stimulus on the unmyelinated
fiber membrane changes its permeability to
Nа
+
ions, a great number of which flow
inside the fiber. In this area the charge of
the membrane changes (the internal
becomes positive, the external — negative).
It generates circular currents (movements
of charged particles) from «+» to «–» along
the whole fiber (Figure 3.9., «а»).

Features of signal conduction along
unmyelinated fibers:
Signals go continuously and the whole fiber
is seized with excitation.
Signals go at a low velocity.
Along unmyelinated fibers signals go to the
internal organs from the nerve centers. However,
the low velocity of the signals and their fading are
not always beneficiary to the human body. That is
why nature made an additional mechanism: the
conduction of signals along myelinated fibers.

Figure — The mechanism of signal
conduction in unmyelinated fibers

The mechanism of signal conduction
in myelinated fibers (Figure). The action
of the threshold stimulus on the membrane
of myelinated fibers at the Ranvier’s node
changes the permeability to Nа
+
ions,
which go inside the fiber. In this part the
charge of the membrane changes, which
also generates circular currents. These
currents go through the intercellular fluid to
the adjacent node, where the charge
changes again. Thus, the excitation
transmits from one part to another. The
reverse movement of the signal is
impossible, as the part through which it has
passed, is at the absolute refractory phase.

Figure — The mechanism of signal
conduction in myelinated fibers

Thus, in myelinated fibers action potentials
occur only at the nodes of Ranvier. The action
potentials are conducted from node to node, and
this is called saltatory conduction. Saltatory
conduction is important for two reasons. Firstly,
this mechanism increases the velocity of
transmission of nerve impulses. Secondly,
saltatory conduction conserves energy for the
axon because only the nodes depolarize,
therefore requiring less energy for re-establishing
the difference between the sodium and
potassium concentrations across the membrane
after a series of nerve impulses.

Features of signal conduction along
myelinated fibers:
Signals go in intermittent motion
(saltatory conduction).
Signals go at a high velocity.
In myelinated fibers signals are
transmitted from analyzers to the СNS,
skeletal muscles i.e. where a high speed of
responses is required.

4. Parabiosis
The scientist N.E.Vvedensky proved that a
part of a nerve changes its lability under the
effect of an alterant (irritant). This happens
due to the fact that excitation lasts longer
within this part and, therefore, at a certain
stage of the alteration, the excitation is not
transmitted through the nerve.
The condition of low lability, i.e.
damage of the normal vital activity of the
nerve is called parabiosis. Parabiosis can be
observed under the action of narcotics, cold or
heat, under the influence of currents and other
stimuli.

The phenomenon of parabiosis was
studied on the example of a nerve-muscle
specimen which consisted of nervous
cells, nerve fibers, and muscles, which
reflected all the changes happening in
nerve fibers (Figure). During the
experiment, a stimulus (for example,
some narcotic on cotton wool) was
applied on some part of nerve fiber.
Through this part the stimulus
transferred and some changes could be
observed.

Figure — The scheme of the nerve-muscle specimen
of parabiosis
A,B,C — electrodes: A — experimental, B — control,
C — in the area of the alteration influence

As a result of the experiment, 3 phases (Figure) of
parabiosis were detected:
1. Provisional or equalizing phase. If to irritate
nerve fiber with stimuli of various strength (weak and
strong), the response of the muscle will always be
identical.
2. If the narcotic continues its action, there comes
the second phase — paradoxical. In this case strong
stimuli induce weak responses, and, on the contrary,
weak stimuli —strong ones.
3. If the effect of the narcotic is not stopped, neither
strong nor weak stimuli can induce a response. This
stage is called inhibitory.
Then, if to terminate the effect of the narcotic and to
wash the damaged part of the nerve, its properties are
recovered in the inverse sequence.

FIGURE — PHASES OF PARABIOSIS
NOTES: THE ARROWS ABOVE SHOW THE STRENGTH OF THE RESPONSE (THE
FORCE OF THE MUSCLE CONTRACTION). THE ARROWS BELOW SHOW THE
STRENGTH OF THE STIMULUS (CURRENT)

5. PHYSIOLOGY OF SYNAPSES

Synapses. Structure of synapses
Synapses are specialized structures which
provide the transmission of excitation from
one neuron to another or to target effector
cells.
The functional role of synapses:
1. They provide functional contacts between
nerves and organs.
2. They promote the regulatory activity of the
СNS.
3. They have plasticity (the amount of signal
which passes through a synapse can change, which
is of an important functional value).
4. They participate in the formation of memory.

The structure of chemical synapses
Nerve fibers approaching a cell form a thickening which
contacts with the cell. This part is the presynaptic
membrane. The opposite membrane is postsynaptic.
Between them there is a cleft filled with a plasma-like fluid.
In the presynaptic terminal, there are neuromediators,
which are capable to excite or inhibit the innervated cell
(Figure).
Myelinated nerve fibers approaching skeletal muscles
make fanlike branchings into end fibers (terminals). The
area of the synapse formation between the nerve
terminations and muscles is called the motor end plate.
The postsynaptic membrane of muscle fibers is thicker and
forms regular folds which increase the surface area of the
postsynaptic membrane. Therefore, a big amount of the
mediator may contact the postsynaptic membrane of
muscle fiber.

FIGURE — STRUCTURE OF THE SYNAPSE

NEUROMUSCULAR
SYNAPSE IS THE
SYNAPSE BETWEEN
MOTOR NEURON
AND
SKELETAL MUSCLE
CELL.
NEUROTRANSMITTER –
ACETYLCHOLINE
Figure — Neuromuscular
synapse

Classification of synapses
1. By the location:
a) peripheral: neuro-muscular, neuro-
secretory, receptor-neuronal;
b) central: axoaxonic, axosomatic,
axodendritic, dendrodendritic,
somatodendritic (Figure).
2. By the effect:
а) excitants;
b) inhibitors.

3. By the mechanisms of signal
conduction:
a) chemical;
b) electrical. They conduct excitation without
participation of the mediator at a high speed
and have bilateral signal conduction. The
structural basis of electrical synapses is the
nexus. These synapses are located in the
endocrine glands, epithelial tissue, СNS, and
heart.
c) mixed.
In some organs excitation can be transmitted
both through chemical and electrical synapses.

FIGURE — ELECTRICAL & CHEMICAL SYNAPSES

4. By the type of the secreted mediator, chemical
synapses are classified into:
а) adrenergic (the mediator is noradrenalin);
b) cholinergic (the mediator is acetylcholine);
c) serotoninergic;
d) glycinergic and others.

Figure — Classification of synapses

Chemical synapses have some common properties:
Excitation in synapses is transmitted only in one
direction. This is provided by the structure of synapses:
the mediator is released only from the presynaptic part
and it interacts with the receptors of the postsynaptic
membrane.
The transmission of excitation in synapses is slower than
that in nerve fibers (synaptic delay).
Excitation is transmitted with the help of special chemical
substances – mediators (neurotransmitters).
In synapses the transformation of the excitation rhythm
occurs.
Synapses have low lability.
Synapses have rapid fatigability.
Synapses have high sensitivity to chemical substances
(including pharmacological drugs — blockers and
others).

Mechanisms of signal transmission in chemical
synapses (on the example of nerve-muscular
synapses)
1. Release of the mediator into the synaptic cleft.
When APs reach the nerve termination (pre-synaptic
membrane), they generate its depolarization. As a
result, calcium ions go inside the terminal. The
increase of the calcium concentration in the nerve
termination promotes the release of acetylcholine into
the synaptic cleft.
2. Diffusion of the mediator to the postsynaptic
membrane and binding with receptors. The mediator
reaches the postsynaptic membrane and binds with
cholinoreceptors located on the postsynaptic
membrane.

FIGURE — STAGES OF SIGNAL TRANSMISSION IN SYNAPSE
Figure — Stages of signal transmission in the
synapse


3. The occurrence of excitation in muscle
fiber. As a result of the interaction of acetylcholine
with the receptors, sodium ions go through the
postsynaptic membrane into the cell and depolarize
the membrane (Figure).
If the initial level of the RMP is — 85 mV, it can
decrease to 10 mV, i.e. partial depolarization
occurs, the excitation does not go further, it stays
in the synapse. These mechanisms cause a synaptic
delay, which may last 0.2–1 ms. Partial
depolarization of the postsynaptic membrane is
called an excitatory postsynaptic potential
(EPP).
Influenced by the EPP in the next part of the
membrane of muscle fiber, there arises a
propagating AP, which produces a muscle
contraction.

In synapses the mediator depending on the
chemical structure can cause depolarization of the
postsynaptic membrane (the excitatory postsynaptic
potential is formed, which provides the exciting effect)
or hyperpolarization of the postsynaptic membrane
(the inhibitory postsynaptic potential is formed, which
provides the inhibitory effect).
4. The removal of acetylcholine from the
synaptic cleft. The enzyme acetylcholinesterase is
located on the external surface of the postsynaptic
membrane. This enzyme disintegrates acetylcholine
and inactivates it.
Some poisons and toxins like botulin can block the
conduction of signals through synapses. For example,
the poison curare contacts the receptors of the
postsynaptic membrane and interferes their interaction
with acetylcholine.

6. Perception of outside stimuli (reception)
Receptors are specific formations which transform
energy of a stimulus into an electrochemical potential
and then into the form of nervous excitation.
Classification of receptors.
By the character of sensations:
1) Visual.
2) Auditory.
3) Olfactory.
4) Gustatory.
5) Tactile.
By location:
1) Exteroreceptors — external (acoustic, visual).
2) Interoreceptors — internal (vestibular and
proprioreceptors).

By the character of stimuli:
1) Photoreceptors — (visual).
2) Mechanoreceptors — (touches and pressure).
3) Thermoreceptors — (cold and warmth).
4) Olfactory.
5) Gustatory.
6) Painreceptor.
By the location of stimuli:
1) Distant — (auditory, visual).
2) Contact — (gustatory, temperature, receptors of
pressure).
All the receptors have adaptation. Adaptation is
decreasing sensitivity to the long effect of a stimulus.

Transformation of stimulus energy
As a result of the interaction of a stimulus and
the receptor membrane, a receptor potential
(RP) appears. How does it happen?
During the contact of the stimulus with the
receptor membrane there is an increase of the
permeability of the membrane to sodium ions
and they get into the sensory terminal, which is
depolarized, and a RP is formed.
The initial conversion of the stimulus into the
RP is called transformation.

The RP excites the initial segment of the sensory
nerve generating a nervous impulse. The frequency
of nervous impulses depends on the RP amplitude.
There are primary-sensitive receptors, which
represent the endings of sensory nerves, and
secondary-sensitive receptors — separate cells
which receive stimulation. These cells are in contact
with the endings of sensory nerves. From these cells
the mediator is released and this results in the
formation of a nervous impulse. A set of receptors
that cause excitation of their own neurons is called
the receptive field; and the areas of the
concentration of receptors belonging to certain
sensory systems are called the reflexogenic zones.
